RNA的二级结构
tRNA的二级结构
tRNA分子只由一条链组成,含有73~94个核苷酸,其中有不少是修饰的核苷酸或修饰的碱基(如次黄嘌吟、硫尿嗑咤和假尿苷等),链内的大多数碱基通过氢键相连,但几乎所有的tRNA分子上不变的核苷酸都位于三叶草结构上的非氢键区域。
构成tRNA二级结构的要素有:环、茎和臂。一个典型tRNA的二级结构像三叶草,含有4个环和4个茎。
环是由链内没有配对的碱基突出而成,茎则是链内互补的碱基之间配对形成的局部A型双螺旋,臂则是紧靠着茎又不属于环的非配对核苷酸。
按照从5'→3'的顺序,4个环依次是D环、反密码子环、可变环和TΨC环。4个茎依次是受体茎、D茎、反密码子茎和TΨC茎。臂有D臂、反密码子臂、TΨC臂和氨基酸臂。
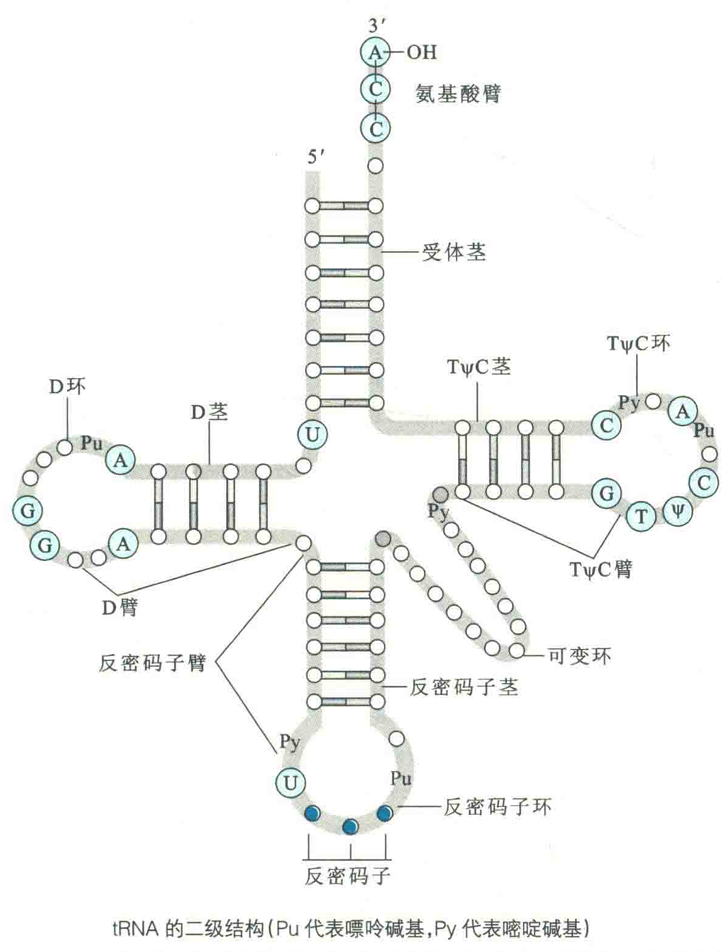
受体茎
受体茎是紧靠氨基酸与tRNA连接形成氨酰tRNA的地方,它由RNA靠近两端的互补序列配对而成。虽然不同的tRNA在一级结构上差别很大,但它们的二级结构却惊人地相似,并且3'端的最后3个核苷酸总是CCA,与3'端的第4个核苷酸一起,并不参与形成受体茎,而是构成接受氨基酸的臂。细胞在合成氨酰tRNA的时候,氨基酸最后被添加到CCA末端腺苷酸的3'-OH上,将来在蛋白质生物合成中作为氨基酸残基的供体。
D环
D环得名于它常常含有一个修饰的碱基,即二氢尿嘧啶(D)。
反密码子环
反密码子环因含有反密码子而得名。反密码子是由三个核苷酸组成的单位,它能够按照碱基互补配对的原则去阅读mRNA上同样由三个核苷酸组成的特定的密码子,从而将核苷酸序列翻译成氨基酸序列。
紧靠反密码子5'端的核苷酸总是U,紧靠3'端的核苷酸总是嘌呤核苷酸(通常烷基化)。
附加环
在反密码子茎和TWC茎之间通常还有一个可变环或附加环,其长度在不同的tRNA分子上会有变化,因此有时可以用它来区分不同的tRNA。
TWC环含有7个没有配对的碱基,包括TΨC序列,其中Ψ即假尿苷。核糖体与tRNA的结合依赖于其对TΨC环的识别。
rRNA的二级结构
核糖体是蛋白质生物合成的场所,其中的RNA被称为核糖体RNA(rRNA)。根据沉降系数的大小,rRNA可分为几种不同的类型。
原核生物有5S rRNA、6S rRNA和23S rRNA;真核生物有5S rRNA、5.8S rRN、8S rRNA和28S rRNA。
在所有的rRNA分子上都发现有大量链内互补的序列,这些序列通过互补配对,使rRNA高度折叠。在不同物种的同一类型的rRNA上存在十分保守的折叠样式。
以16S rRNA为例,其内部存在大量的短螺旋,螺旋之间夹杂着多种形式的环和突起,这是根据形成氢键的序列比对出来的。比较不同物种来源的16S rRNA的一级结构和二级结构发现,尽管它们在一级结构上相似性并不高,但它们的二级结构也惊人地相似。
显然,16S rRNA的分子进化是二级结构在起作用,而不是表现在核苷酸的序列上。换句话说,16S rRNA在进化中,只要保证它的二级结构基本不变就行,而不用在乎其一级结构的变化。
再以5s rRNA为例,它的二级结构由5段螺旋(I~V)、1个突环(环A)、2个发夹环(环C和环E)、2个内部环(环B和环E)和一个铰链区组成。
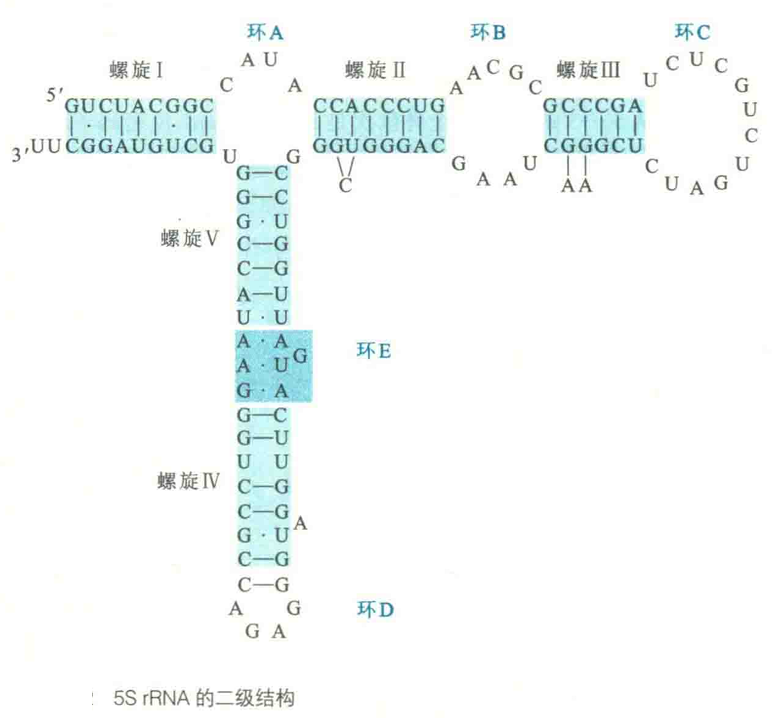
图中的内部环E并没有画成环,是因为该环的5'端序列和3'端序列在三级结构上通过非Watson-Crick碱基对结合在一起。
mRNA的二级结构
mRNA的种类繁多,对各种mRNA的二级结构的研究却很少。
实际上,人们关心更多的是它们的一级结构,这是因为编码多肽或蛋白质氨基酸序列的是其一级结构。
然而,研究发现,mRNA分子的二级结构,尤其是两端的二级结构,对翻译有一定的影响,某些mRNA正是借助于末端特殊的二级结构对基因的表达进行调控。
出现在mRNA分子上最多的二级结构部件也是茎环结构。
DNA-RNA杂交双链的二级结构
由DNA和RNA形成的杂交双链形成的是A型双螺旋。这样的结构会在体内DNA复制、转录和反转录(也称逆转录)的时候出现。
在DNA复制的时候,先要合成一段短的RNA引物,这段RNA序列与DNA一条链上的部分序列是互补的,它们之间形成局部的A型双螺旋;在基因转录的时候,DNA一条链上的一段核苷酸序列作为模板,合成各种RNA分子,被转录的RNA分子总是有7~8个核苷酸与DNA的模板链以A型双螺旋结合;而在反转录的时候,RNA是模板,合成的cDNA与其也是形成杂交双螺旋结构。